| French
|
Arabic
|
English
|
| Enzymes allostériques, Enzymes régulatrices | إنزيمات الموقع الآخر، إنزيمات ألوستيرية، إنزيمات منظمة | Allosteric enzymes, Regulatory enzymes |
Les enzymes allostériques (إنزيمات الموقع الآخر - إنزيمات ألوستيرية, Allosteric enzymes) assurent la régulation des chaînes métaboliques. Cette faculté est due à la présence d'un site régulateur (en plus du site actif) qui réagit avec des modulateurs qui viennent d'ailleurs (allosterie, Allostery). Les enzymes allostériques sont des enzymes assurant la régulation des chaînes métaboliques.
Propriétés des enzymes allostériques
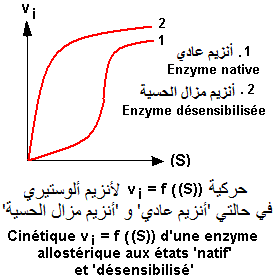
Comportement cinétique des enzymes allostériques.
Les enzymes allostériques (E) sont des enzymes de régulation des chaînes métaboliques. Elles sont caractérisées par des comportements cinétiques différents de ceux des enzymes michaelliennes. Contrairement à ces dernières (cinétique 'HYPERBOLIQUE'), les enzymes allostérique présentent une cinétique 'SIGMOIDE'

Effet
allostérique: le terme d'allostérie a été
utilisé au départ pour expliquer l'inhibition
compétitive de certaines enzymes par des molécules ne présentant pas ou peu de ressemblance au substrat initial.
Dans le type classique d'inhibition compétitive, l'inhibiteur (I) est un analogue structural du substrat (S) et tend à occuper le même site que ce dernier. Il s'agit d'un inhibiteur isostérique
(isostérie) (voir Fig). Par contre, l'inhibiteur ne montrant pas une analogie structurale avec le substrat occupera un autre
site sur l'enzyme. Il est qualifié d'inhibiteur allostérique
(allostérie). Ces différentes interactions permettent
de concevoir l'existence de plusieurs formes d'enzyme et la présence
de site allostérique et de site actif.

Effet de coopérativité: la coopérativité traduit le fait que la fixation sur l'enzyme d'une molécule d'un effecteur allostérique (exemples: activateur allostérique (A) et inhibiteur allostérique (I)) influe sur la fixation des molécules suivantes.
Dans le cas de coopérativité en présence du substrat (sigmïde de v=f((S)), Si l'effecteur allostérique est le substrat lui même on parle de modulation homotrope. Si l'effecteur est différent du substrat on parle de modulation hétérotrope. Dans une coopérativité positive, une molécule d'un effecteur entraîne l'augmentation de l'affinité pour les mêmes molécules et vice versa pour une coopérativité négative.
Le traitement d'une enzyme allostérique par des agents physiques (ex.chauffage) ou chimiques (urée, dérivés mercuriques) s'accompagne d'une perte de la sensibilité de l'enzyme aux effecteurs allostériques (désensibilisation). Cependant, l'activité enzymatique persiste. Seul, le site allostérique est détruit. Il en résulte une perte du phénomène de coopérativité et la cinétique devient hyperbolique.
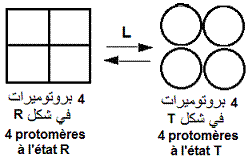
Nature oligomérique des enzymes allostériques et transition allostérique R <---> T.
l'enzyme allostérique est un ensemble de sous-unités spécifiques associées. Celles ci coopèrent en vue d'une même fonction. C'est une structure quaternaire.
Transition d'aggrégation-désagrégation:
Les premières idées faites sur l'activité des enzymes allostériques (1960) considéraient que l'état
fonctionnel d'une enzyme allostérique était un état d'aggrégation. L'état de désagrégation
correspondait à l'état inactif (transition d'aggrégation-désagrégation)
(Fig). Cette hypothèse est inéxacte in vivo.
Transition R <---> T:
La transition allostérique modifie les
forces de liaisons qui associent les sou-unités entre elles sans aller à l'état de dissociation. La molécule
entière se trouve soit dans un état contraint (T)
soit dans un état relaché (R). Cette transformation de la molécule est réversible.
Les différentes sous-unités (protomères) d'une enzyme allostérique sont caractérisées par: l'existence
d'un centre de symetrie pour les protomères associés, la présence au sein de chaque protomère d'un seul site
stéréospécifique de chaque effecteur et la conservation
de la symetrie de la molécule entière malgré
les changements conformationnels.
Les deux formes R et T peuvent être actives l'une et l'autre. Leur rapport change de manière complexe en fonction des différents
effecteurs. Il a été noté que les formes R et T de l'Aspartate transcarbamylase n'ont pas le même pH optimum
(pH = 8,2 pour la forme R et pH = 6,8 pour la forme T). Elles diffèrent
également par leur spectre aux rayons X.
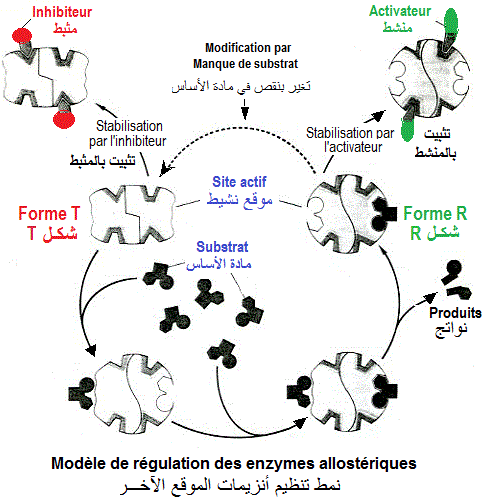
Ci contre le modèle de coopérativité positive par le substrat pour une enzyme allostérique. L'enzyme est d'abord,
présente sous la forme T (moins active). La fixation d'une molécule de substrat (S) sur le site spécifique de la
forme T, induit un changement conformationnel du site actif encore
libre le rendant plus favorable à fixer plus de S. L'enzyme
devient sous forme R (plus active) maintenue aussi lontemps que la
quantité de S est suffisante pour occuper le site actif.

Ci contre, un modèle de la régulation d'une enzyme allostérique par les modulateurs de type 'inhibiteur allostérique' (effecteur négatif) et 'activateur allostérique'(effecteur positif). La forme T de l'enzyme, moins active, est transformée en forme R, plus active, par fixation du substrat. La forme R, très active, induite par la fixation de S, est stabilisée par un activateur. La fixation de l'inhibiteur sur l'enzyme s'accompagne d'un rendement faible en activité enzymatique.
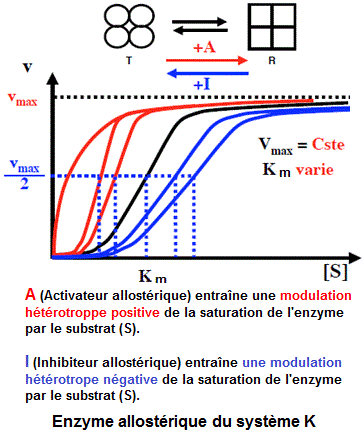
Modulations allostériques de types K et V
Trois types d'enzymes allostériques peuvent être distingués selon les effets exercés sur elles
par les effecteurs homotropes et hétérotropes qui les concernent. Ainsi, on distigue:
- Les enzymes du système K où l'effecteur ne modifie que l'affinité apparente (relative à Km) de l'enzyme
pour le substrat.
- Les enzymes du système V où l'effecteur ne modifie que la vitesse maximale (Vmax) de la réaction.
- Les enzymes du système mixte où l'effecteur ne modifie les deux paramètres Km et Vmax.
Ce sont des enzymes où le substrat et l'effecteur présentent, tous, des affinités différentes pour les formes R et T de l'enzyme. Il en résulte une fixation toujours sigmoïde.

Les formes R et T ont une même vitesse maximale. Sur le plan cinétique, ce sont des enzymes où l'effecteur ne peut modifier que l'affinité apparente (relative à Km) de l'enzyme pour le substrat. L'affinité pour le substrat (S) diminue en présence d'un inhibiteur allostérique (I). Elle augmente en présence d'un activateur allostérique (A).
 |
 |
 |
 |
Enzymes allostériques du système K (suite).
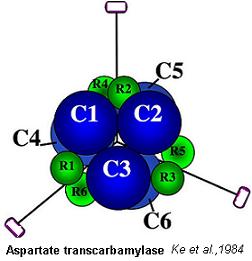
Exemples d'enzymes allostériques du système K: Phosphofructokinase (PFK), Thréonine désaminase, Aspartate transcarbamylase, (voir structure oligomérique de l'Aspartate transcarbamylase)
Voir ..... Aspartate transcarbamylase. Examen
Le CTP, produit de la voie des nucléotides pyrimidiques, est un inhibiteur allostérique.
L'ATP, produit de la voie des nucléotides puriques, est un effecteur activateur allostérique.
Le CTP et l'ATP sont impliqués dans la régulation. L'enzyme aspartate transcarbamylase est donc régulée par le rapport ATP/CTP de la cellule.
Enzymes allostériques du système V (plus rares).
Les enzymes allostériques du système V sont caractérisées par le fait que le substrat (S) a la même affinité pour les deux formes d'enzyme R et T. Tout se passe comme s'il y'a une seule forme d'enzyme. Il en résulte une fixation hyperbolique de S. La coopérativité homotrope
est absente. Néanmoins, les effecteurs A et I ont des affinités différentes pour chaque forme R et T. Il en résulte
une fixation sigmoïde témoignant d'une coopérativité
homotrope pour A et I.
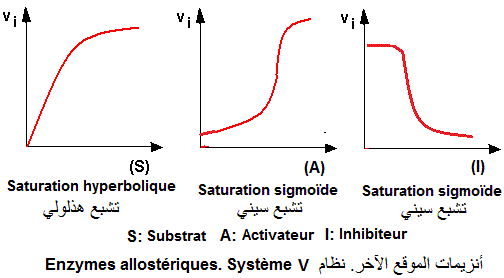
Les formes R et T sont différentes dans leurs activités catalytiques (VmaxR (forme R) différent de VmaxT(forme T)). En se fixant sur l'une des formes, un effecteur va augmenter ou diminuer Vmax par simple déplacement de l'équilibre R <---> T. Si R est catalytiquement plus active la fixation d'un activateur sur cette forme entraîne le déplacement de l'équilibre vers cette forme. Il en résulte une augmentation de Vmax. Le raisonnement inverse est valable pour un inhibiteur allostérique se fixant sur la forme T.

-
Interprétations moléculaires des modulations allostériques.
- Modèle de coopérativité de Hill (1909).
- Modèle de Monod, Wymann et Changeux (MWC).
- Modèle de Koshland
Liens utiles:
- Allosteric enzymes (Ar, Eng)
- Enzymes allostériques. Exercices
- Enzymes et Métabolisme. Exercice
- Régulation enzymatique du métabolisme
التنظيم الإنزيمي للاستقلاب
- Enzymologie-enzymes
(exercices)
- Comparaison Hexokinase et Glucokinase. Exercice
- QCM-Enzymes. Structure et fonction (bases)
-
Hexokinase
- Coenzyme
Atransférase
- Amine
oxydase
- Inhibiteurs.
Applications
- QCM-Inhibiteurs-1
- QCM-Inhibiteurs-2.
- Enzymologie. Examen-1
- Enzymologie. Examen-2
- Enzymologie. Examen-3
- Enzymologie. Examen 4
- Enzymologie. Examen 5
- Polyphénoloxydase. Examen S5
--> Retour à SOMMAIRE
PASSER UN CONTROLE NOTE ET COMMENTE SUR L'ENZYMOLOGIE

Ouvrages de la préparation de la transition Secondaire-Supérieur


-
من علوم الحياة إلى البيوكيمياء
Des Sciences de la vie au Lycée à la Biochimie à l'Université
-
تحميل ملفات في مواضيع البيوكيمياء
Téléchargement de fichiers en Biochimie

BIOCHIMIE. TP CONSIGNES (Vidéos)
Chaine
Youtube (abonnement). Plusieurs vidéos multilingues

مجلة إخبارية - Newsletter

ANNONCES, INFORMATIONS
Congrès, Conférences,...
- Congrès, Conférences
- Cours, Workshops
- Bourses, Postdocs
- Biotech-ecolo annonces
CONCOURS:
Concours de
Pharmacie
Afin de pouvoir continuer à servir les visiteurs, soutenez nos actions sur le site takween en faisant acquisition des ouvrages et supports pédagogiques desitinés à améliorer l'enseignement et la recherche scientfique en Biochimie.


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com
{kind=link}
{kind=link}
{kind=link}