
Après avoir procédé à l'extraction et la purification de l'ADN des plantes (Première partie du TP S5), (Pour rappel, la purification de l'ADN et sa quantification ont fait l'objet de TP précédents; en S3). Il y'a lieu de passer à son analyse par électrophorèse sur gel d'agarose. Les résultats des traitements de l'ADN (hydrolyse par enzyme de restriction, amplification par PCR) sont tributaire du degré de pureté de l'ADN et de son état intact ou altéré au cours des différentes étapes de la purification.
L'électrophorèse
reste l'outil le plus important dans l'analyse qualitative (voir même quantitative) des acides nucléiques (ADN, ARN). Son principe doit être bien compris. Les critères de séparation utilisés sont la
taille (poids moléculaire) et l'ionicité
(charge électrique). Comme les acides nucléiques
sont chargés négativement (grâce au phosphate),
le critère de la charge électrique n'est pas comptabilisé
dans la séparation (toutes les molécules migrent dans
la même direction, +) et les fragments de l'ADN sont séparés
uniquement en fonction du poids moléculaire. L'analyse des
acides nucléiques (en particulier l'ADN) par électrophorèse consiste d'abord à lui faire subir un traitement préalable
dont 1/ coupure (restriction) par une ou plusieurs enzymes de restriction
(technique RFLP) ou 2/ amplification de certains fragments (ou régions) par la réaction
de polymérisation en chaîne (PCR, polymerisation chain
reaction). En plus des critères de l'ionicité et
de la taille, la forme
des molécules intervient dans le cas des plasmides
(voir ci dessous). De ce fait, l'implication
de la forme des plasmides dans l'électrophorèse complique
la détermination exacte de leurs tailles.
La complexité d'un génome et sa grande taille rendent
impossible l'analyse directe des profils de restriction de l'ADN des
eucaryotes. Ceci est dû au nombre élevé des fragments
de restriction obtenus après action d'une enzyme de restriction
(supérieur à 1000000). Dans la majorité des cas,
le profil de restriction obtenu après révélation (ou coloration) de l'ADN montre un voile (ou continuum, smear)
représentant tous les fragments de restriction tassés sur le gel (tailles faibles vers +) et difficiles à distinguer. Ce
problème est résolu par recours à des techniques supplémentaires qui permettent de mettre en évidence
uniquement les fragments interessant l'expérimentateur. Ceci
est rendu possible par l'utilisation d'une sonde (probe, مجس)
marquée préparée pour réagir spécifiquement
(par hybridation) avec des séquences précises à travers la technique de transfert (southern blotting).
Pour réaliser cette tâche, il faut effectuer un transfert
des bandes d'ADN du gel sur une membrane de nitrocellulose (ou nylon).
Une reconstitution d'un film, de l'extraction
de l'ADN à son analyse, existe sur CD.
L'éléctrophorèse peut être réalisée horizontalement ou verticalement (avec les gels de polyacrylamide).
Electrophorèse des plasmides. Effet de la forme d'ADN
Les
plasmides comme pBr 322, existent sous 3 formes dont l'une peut résulter
de l'autre suite à des altérations ou une longue conservations:
- Forme superenroulée montrée
par le plasmide bien conservé (supercoiled form, en Anglais).
- Forme relachée ou forme circulaire
ouverte (Open circular, nicked circle, en Anglais) se rencontrant lors d'une mauvaise conservation du plasmide et portant une coupure
sur un seul brin.
- Forme linéaire (Linear form,
en Anglais), pouvant résulter de la digestion du plasmide par
les enzymes de restriction. Elle correspond à une coupure sur
les deux brins.
Par électrophorèse sur gel d'agarose, on peut révéler
les formes superenroulées et relâchées en même temps (voir schéma). La conformation de l'ADN affecte la migration
électrophorétique. Pour supprimer cet effet de la forme,
le poids moléculaire est déterminé sur la base de l'ADN linéaire (résultant de la restriction
ou de l'amplification par PCR). Le schéma ci dessous montré
le résultat d'une digestion progressive d'un plasmide comme
révélé par électrophorèse.

Préparation des échantillons pour leur séparation par électrophorèse
Tous les échantillons devant êtranalysés, doivent être mélangés à un Tampon de charge (loading buffer, منظم التحميل) qui les rends visibles (bleu de couleur)et denses (contenant glycérol):

- tampon TBE (pH 8,3)
- Xylène cyanol 0,02% -> migre en prallèle avec ADN bicaténaire de 4000 pb sur les gels d'agarose 1%.
- Bleu de bromophénol 0,02% --> migre en prallèle avec ADN bicaténaire de 500 pb sur les gels d'agarose 1%.
- Orange G --> migre en prallèle avec ADN bicaténaire de 50 pb sur les gels d'agarose 1%.
- Glycérol 3% --> augmente la densité des échantillons d'ADN.
Les 3 colorants (voir formules ci dessous) permettent de suivre la migration des fragments d'ADN.

En plus des échantillons d'ADN purifié, Les dépôts concernent aussi:
- Marqueurs des poids moléculaires (pb) correspondant à 1 Kb DNA ladder (10 fragments)VOIR
L'ELECTROPHOREGRAMME DES MARQUEURS DE PM
- Dilutions successives (préparées dans le tampons de charge, pH 8,3) de 4 préparations d'ADN
de plantes préalablement purifiés (A, B, C et D).
- Plasmide pL9 natif (plasmide recombiné)
- Plasmide pL9 hydrolysé par Hind III (10-100 ng)
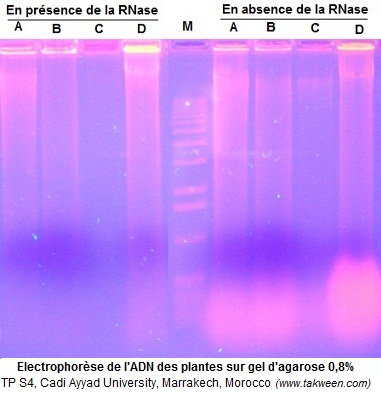
- ADN des plantes (A,B, C et D) hydrolysé par Hind III en présence et en absence de la RNase.
Les plasmides recombinés sont utiles dans les opérations du génie génétique et des biotechnologies modernes.


Le
support de migration le plus utilisé dans l'éléctrophorèse
de l'ADN est constitué de l'agarose (0,5-2,5%, environ) suivi
des gels de polyacrylamides. Les étudiants déposenet
de faibles volume d'ADN hydrolysé par une enzyme de restriction
comme Hind III.
Un support de migration constitué d'un gel d'agarose à
0,8% qui permet de séparer des frangment
de tailles comprise entre 800 et 10000 pb, environ (QCM
sur choix du gel?).

-
Peser l'agarose (polymère à base d'agar purifié) dans un erlenmeyer, en fonction du pourcentage choisi et du volume de gel à préparer.
- Placer l'erlenmeyer contenant la solution d'agarose dans un bain Marie bouillant pendant au moins 20 minutes en agitant de temps en temps.
- Lorsque l'agarose est bien fondue (solution limpide), retirer l'erlenmeyer et laisser refroidir jusqu'à 60°C, environ (voir la vidéo ci dessus, laboratoire BBP, FSSM, Marrakech, Morocco).
- Remarque: si on opte pour la révélation de l'ADN par le Bromure d'éthidium (BET), il sera possible d'ajouter dans la solution d'agarose un volume précis du BET pour avoir une concentration finale de 0.5 µg/ml. Autrement le BET est ajouté en solution après l'électrophorèse.
- Couler le gel d'agarose dans le moule préalablement préparé. Eliminer les bulles. Placer les peignes (servant pour ménager des puits dans le gel) vers une extrêmité du gel et laisser refroidir au moins 45 minutes.



Qualité du dépôt :1) pas de bulles, 2)échantillon totalement contenu dans le puits, 3)pas de débordements, 4)pas de puits percé.
- Avant de lancer l'électrophorèse, enlever les cales et les peignes délicatement pour ne pas altérer les puits.
Réglage du voltage:
La séparation se fait à voltage constant à raison de 100 volt pendant environ 2 heures de migration (l'intensité en mA reste variable). Normalement, un champ électrique de 5 V/cm est appliqué au gel. La distance entre les électrodes permet de calculer le voltage à appliquer aux bornes du générateur.
Par exemple : si la distance entre les électrodes fait 12 cm, le voltage à régler est de 5 x 12, soit 60 V.
- Un champs plus important accélère proportionnellement la migration, limite la diffusion, mais fait chauffer le gel par effet Joules.
- Si on veut accélérer la migration, il faut augmenter le voltage, mais baisser la force ionique du tampon pour diminuer l'effet Joules (voir QCM électrophorèse).
Rappel des relations liant le voltage, la force ionique et l'effet Joules :
- La relation entre le courant I, la tension V et la résistance R est exprimée comme dans la loi d'Ohm : V=R.I
L'augmentation de la tension V et l'augmentation correspondante du courant I provoqueront l'un des principaux problèmes posés dans l'électrophorèse, en l'occurrence
la génération de chaleur (chauffage). Ceci peut être illustré par l'équation suivante où la puissance du courant P (mesuré en watts) généré
durant l'électrophorèse est égal au produit de la résistance multiplié par le carré du courant
: P = V.I=RI²(correspond au chauffage).
- rho = 1 / R = K.Fi : la conductance (rho) est proportionnelle à la force ionique (Fi) et inversement proportionnelle à la la résistance.
- A voltage constant: si l'on baisse la force ionique, la conductance diminue, donc la résistance augmente, I diminue (car I = V/R) et aussi P diminue (car P =VI): donc chauffage diminue
Conclusion : à voltage constant, diluer le tampon permet de diminuer l'échauffement du gel. On peut donc se permettre d'augmenter
la vitesse de migration, en augmentant le voltage à condition de diminuer la force ionique du tampon (par exemple passer du tampon TBE 1 X à 0,5X).
Remarque: Le tampon TAE (Tris-Acétate-EDTA, pH 8,0), de conductivité
élvée, génère plus de chauffage que les tampons de faible force ionique comme TBE.
La révélation de l'ADN sur le gel d'agarose est réalisée par le bromure d'éthidum qui est ajouté dans le gel d'agarose à l'état liquide (avant qu'il soit solide) ou simplement en solution dans laquelle on fait incuber le gel pendant 20 minutes, environ, une fois l'éléctrophorèse est terminée. La concentration finale du BET de 0.5 µg/ml.
Les fragments d'ADN sont rendus visibles par exposition du gel aux rayons UV à l'obscurité. Lorsqu'il est ajouté
dans le gel, il peut causer un ralentissement de la migration d'environ
15%.
Le bromure d'éthidium se lie à l'ADN bicaténaire par intercalation (= agent intercalent).

Les rayons UV sont dangereux pour les yeux : il faut porter des lunettes, ou disposer d'une enceinte dans laquelle sont placés le transilluminateur et une caméra.

Pour déterminer le poids moléculaires des fragment d'ADN séparés sur le gel d'agarose, les étudiants tracent la courbe du logarithme des poids moléculaires de fragments d'ADN de poids connus (marqueurs de PM, DNA-ladder) en fonction de la distance parcourue (ou aussi la mobilité relaive) par les différents fragments. La droite : log (taille) = f (distance de migration) permet de déterminer la taille en paires de base d'un fragment d'ADN inconnu.---> VOIR L'ELECTROPHOREGRAMME DES MARQUEURS DE PM

Estimation de la quantité d'ADN contenu dans les extraits d'ADN des plantes.
Comme exemple, prenons le profil électrophorétique du plasmide hydrolysé avec Hind III et dont la quantité déposée est 60 ng (profil 26 de l'électrophorégramme). Le profil montre deux bandes correspondant à l'insert(voir structure du plasmide) de petite taille (environ 1000 pb, d'après la courbe des PM) et le vecteur de grande taille migrant moins vite par rapport à l'insert et présente environ 3000 pb, comme taille (d'après la courbe log (PM) = f(distance parcourue)). La taille totale du plasmide sera donc 1000 + 3000 = 4000 pb. Elle correspond à 60 ng. Donc, l'insert (1000 pb) correspond à 15 ng et le vecteur correspond à 45 ng. Si l'estimation de l'intensité de l'ADN dilué par exemple 5 fois, est proche de l'intensité de la bande du vecteur (45 ng)on pourra déduire que la quantité d'ADN dans 10 µl de l'extrait d'ADN dilué déposé est également 45 ng. Comme l'extrait de départ a été dilué 5 fois, la quantité d'ADN sera donc (45 x 5) g. Si cette quantité était dans une prise d'essai de 8 microlitres d'ADN , la quantité d'ADN dans 50 microlitres sera 45 x 5 x 50/8 = 1406,25 ng.
HindIII est une enzyme de restriction isolée de Haemophilus influenzae. En présence de Mg++, elle clive l'ADN au niveau de la séquence AAGCTT.
5'-A |A G C T T-3'
3'-T T C G A| A-5' .
QUESTION. L'ADN de plantes purifié suivant un protocole précis a été soumis à une hydrolyse par HindIII en présence ou en absence de la RNase puis séparé par éléctrophorèse sur gel d'agarose 0,8%. Sur l'électrophorégramme suivant, où se trouve l'ARN? - cocher la réponse juste
-
(!Bande intense en haut du gel pour les extraits d'ADN hydrolysés par RNase)(Bande intense et diffuse en bas du gel pour les extraits d'ADN non hydrolysés par RNase)(!Bande intense en haut du gel pour les extraits d'ADN non hydrolysés par RNase)
Electrophorèse de l'ADN (DNA) sur gel d'agarose (Préparation des échantillons, dépôt, migration et révélation par Bromure d'Ethidium, ...)
- Protocole d'extraction et de purification de l'ADN
- QCM purification ADN des plantes (TP S5)
- DNA electrophoresis on agarose gel
- QCM Electrophorèse
- ADN. Structure 3D par JMOL
- Biologie
moléculaire. Examen TP S5
- Technique d'électrophorèse
Extraction
de l'ADN de plasmide, clonage, séquençage. Exercice
- Clonage
des gènes et génie génétique
- Biologie moléculaire.
Examen TP S5 2016
- ADN. Quantification
par spectrophotometrie (TP S3)
Afin de pouvoir continuer à servir les visiteurs, soutenez nos actions sur le site takween en faisant acquisition des ouvrages et supports pédagogiques desitinés à améliorer l'enseignement et la recherche scientfique en Biochimie.


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com
{kind=link}
{kind=link}
{kind=link}