| French | English | Arabic |
| Métabolisme des lipides; acides gras | Lipid metabolism, fatty acids | استقلاب الدهنيات، أحماض دهنية |
L'un des buts du métabolisme des lipides (voir schéma) est de produire l'énergie par béta-oxydation des acides gras provenant de la lipolyse des lipides ou de la nourriture.
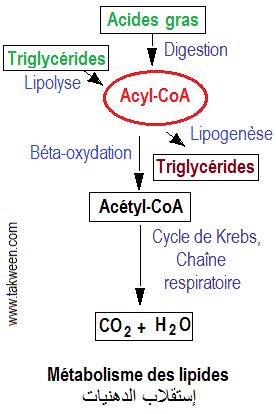
La béta-oxydation des acides gras et sa régulation peuvent être résumées par le schéma suivant:

Le catabolisme des acides gras a été élucidé par des expériences de marquage d'éléménts. Ainsi Lynen a découvert le mécanisme du catabolisme des acides gras dit 'béta-oxydation' qui consiste à la scission deux par deux atomes de carbone d'un acide gras en commençant par la partie carboxyle.

Le catabolisme des acides gras se déroule dans les mitochondries(contrairement à leur biosynthèse, se déroulant dans le cytoplasme). Leur transport du cytoplasme s'effectue sous forme combinée (comme ester de carnitine, par exemple).
1/ Activation des acides gras au niveau de la membrane mitochondriale externe
Après
leur libération par lipolyse dans le cytoplasme, Le catabolisme
des acides gras est réalisé dans les mitochondries, sur
leurs formes métaboliques actives, les acyl-coenzyme
A. L'activation des acides gras se déroule en deux phases;
1/ Formation des acyls CoA et 2/ Transfert des acyls CoA du cytoplasme
vers la matrice de la mitochondrie.
- Formation de l'Acyl-CoA: catalysée par une thiokinase, l'acyl-CoA
synthétase au niveau de la membrane mitochondriale
externe.
R-COOH + ATP + HSCoA <----> R-CO-SCoA + AMP + PP (enzyme: thiokinase)
L'activation est effectuée dans la membrane mitochondriale externe.
Leur dégradation (oxydation) se déroule dans la matrice
mitochondriale (voir schéma).
- Transfert de l'acyl CoA dans la mitochondrie
Les acyls CoA à chaîne courte pénétrent facilement dans la matrice mitochondriale. La membrane mitochondriale interne est imperméable aux acyls Co à chaînes longues (12 C-18C).
Les groupements acyls traversent cette membrane sous forme d'acyl-carnitine.
2/
béta-oxydation des acides gras au niveau de la matrice mitochondriale
Dans la matrice, la scission d'un chaînon dicarboné
à partir de l'extrémité carboxylique de l'acyl
CoA est réalisée en 4 étapes: 1/ Déshydrogénation
des carbones alpha et béta (oxydation par FAD), 2/ Addition
d'une molécule d'eau (hydratation), 3/ Deuxième
déshydrogénation (oxydation par NAD) et 4/ Thiolyse
(coupure par CoA). Cette dernière conduit à un acyl CoA
à (n-2) carbones et libération d'un acétyl-CoA..

Remarque: les étapes 1, 2 et 3 sont très similaires à celles du cycle de Krebs allant du succinate à l'oxaloacétate (Succinate -> Fumarate -> Malate -> Oxaloacétate).
Les acétyl-CoA produits par l'oxydation des acides gras entrent dans le cycle de Krebs. Les FADH2 et les NADH produits sont oxydés par la chaîne mitochondriale de transport des électrons pour permettre la production d'ATP.
3/ Hélice de Lynen
Chaque tour de spire de l'hélice conduit à la perte d'un acétyl-CoA et de 4 atomes d'hydrogène.
Bilan de la béta-oxydation:le bilan de la béta-oxydation de l'acide palmitique est:
| Acide palmitique + ATP + 8 HSCoA + 7 FAD + 7 NAD+ + 8 H2O | --> | 8 CH3CO-SCoA + AMP + 2 Pi + 7 FADH2 + 7 NADH, H+ |
Bilan
énergétique pour l'acide palmitique:
Activation de l'acide gras: - 2ATP
- Réoxydation de 7 FADH2 par la chaîne respiratoire: 7x 2 = 14 ATP
- Réoxydation de 7 NADH, H+ par la chaîne respiratoire: 7x 3 = 21 ATP
- Oxydation de 8 CH3-CO-SCoA par la Cycle de Krebs: 8x 12 = 96
ATP*
Bilan énergétique: (14 + 21 + 96) - 2 = 129 ATP libérées
par oxydation d'une molécule d'acide palmitique.
* L'oxydation d'une molécule d'acétyl-CoA par le cycle de Krebs donne 1 ATP + 3 NADH + 1 FADH2.
 |
 |
 |
 |
 |
Le
catabolisme des lipides (béta-oxydation) ne peut s'effectuer normalement que s'il coexiste avec le catabolisme des glucides. C'est pour cela on dit 'les lipides brulent au feu des glucides - تحترق
الدهنيات بنار السكريات .
En absence de glucides (en cas de diabète et jeûne), l'Acétyl-CoA, utilisé en priorité par le cycle de Krebs (respiration mitochondriale), ne peut malheureusement pas pénétrer dans le cycle de Krebs, à cause du manque d'acide oxaloacétique converti en glucose pour le cerveau (par la voie de la néoglucogenèse, voir schéma). L'acétyl-CoA s'accumule et donne des coprs cétoniques (cétogenèse), comme l'acide acétoacétique, l'acide béta hydroxybutyrique et l'acétone. Le premier corps cétonique formé est l'acéto-acétate. Il est obtenu par condensation de deux acétyl-CoA, suite à trois réactions enzymatiques. Les deux autres corps cétoniques sont obtenus à partir de l'acéto-acétate, grâce à l'activité de deux enzymes distinctes. L'acide béta-Hydroxybutyrique est formé par reduction grâce a une enzyme à NAD+. L'acétone est formée par décarboxylation de l'acétoacétate. les molécules d'Acétyl
CoA accumulées en corps cétoniques, sont libérées
dans le sang et utilisées à des fins énergétiques
par d'autres tissus. Les corps cétoniques sanguins sont
captés par les cellules dans lesquelles le cycle de Krebs fonctionne
à plein régime, comme les cellules du cœur. Ces corps
subissent alors la respiration cellulaire. En cas d'excès important dans le sang, les corps cétoniques deviennent toxiques.
Ils sont excrétés dans les urines ou dans l'air au niveau des poumons où ils provoquent une halitose (mauvaise haleine due à l'accumulation d'acétone dans l'air expiré (odeur proche de celle de la pomme).

------> Voir TP S4 sur la Glucose 6 phosphatase et la néoglucogenèse
Régulation de la cétogenèse
La cétogenèse est régulée par des facteurs métaboliques et des facteurs hormonaux.
- Facteurs métaboliques de la régulation de la cétogenèse
La cétogenèse est augmentée lorsqu'il y'a une diminution de l'utilisation des glucides avec augmentation de celle des acides gras
A- Utilisation exagérée des acides gras libres :
l'augmentation des acides gras au niveau du foie favorise la cétogenèse par:
1/ Modification du potentiel d'oxydo-réduction. Ainsi, l'augmentation de la teneur en NADH avec augmentation du rapport NADH/NAD, conduit à une diminution de l'activité des réactions catalysées par les enzymes NAD dépendantes, en particulier celles de la formation de l'Oxaloacétate à partir du Malate. Ce
qui empêche l'Acétyl CoA d'entrer dans le cycle de Krebs.
2/ Inhibitions enzymatiques directes :
- La citrate synthétase empêche la fixation de l'Acétyl CoA sur l'Oxaloacétate. La diminution du citrate diminue la sortie des acyétyl-CoA de la mitochondrie, ce qui limite la
lipogenèse. Ces inhibitions enzymatiques bloquent l'utilisation de l'acétyl-CoA dans le cycle de Krebs et la lipogenèse, orientant l'acétate vers la formation intra mitochondriale
de corps cétoniques. Aussi, la régulation de la dégradation des acides gras libres: dépend du taux de malonyl CoA, qui conditionne la pénétration intra mitochondriale des acides gras libres et leur oxydation en acétyl-CoA.
B- Diminution de l'utilisation des glucides :
- La carence en glucides entraîne une cétogenèse par 2 mécanismes :
* blocage du cycle de Krebs : par diminution de la formation d'Oxaloacétate.
* inhibition de la lipogenèse : par diminution de l'ATP
formée par la voie Embden Mayerhuff et NADH.
- Facteurs hormonaux de la régulation de la cétogenèse
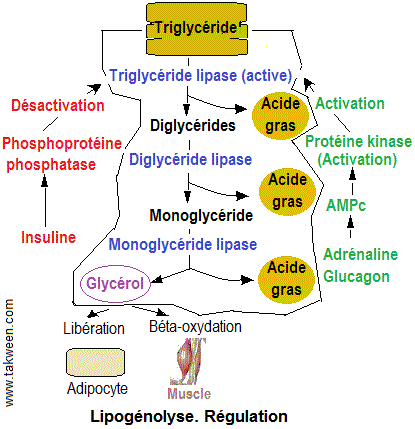
A- Facteurs cétogènes : augmentent l'utilisation
des acides gras et/ou diminuent celle du glucose. On peut citer les
hormones lipolytiques (catécholamines et glucagon activant la
lipase qui libère les Aacides gras à partir des triglycérides) et d'autres hormones.
B- Facteurs anti-cétogènes comme l'insuline qui augmente l'utilisation hépatique du glucose et augmente la pénétration intracellulaire du glucose et son utilisation
au niveau des tissus périphériques. Elle diminue la lipolyse
et augmente al lipogenèse, en favorisant l'entrée
des acides gras libres dans les adipocytes et en favorisant la synthèse
des triglycérides.
LIENS UTILES
-
Métabolisme général
- Glycolysis and gluconeogenesis
regulations
- Homéostasie de
la glycémie
- QCM Métabolisme des
glucides
-
Biochimie métabolique
(métabolisme). Examen S4
-
Glycolyse et production d'ATP
-
TP néoglucogenèse
(Glucose-6 phosphatase)
-
Glycéraldéhyde-3-Phosphate
Déshydrogénase (GAPDH)
-
QCM métabolisme général
-
QCM Chaîne respiratoire
- Comparaison Hexokinase et Glucokinase. Exercice
- QCM Néoglucogenèse (TP Métabolisme, S4)
- تقويم ذاتي مع التوقيت و التنقيط
Contrôle d'autoévaluation, temporisé, noté et commenté


360 pages, 2018, ISBN : 978-9920-35-345-8 + DVD mis à jour + Assistance.
Comment avoir ces livres ? - أين يمكن اقتناء الكتب؟
Chaine
Youtube (abonnement). Vidéos multilingues

-
من علوم الحياة إلى البيوكيمياء
Des Sciences de la vie au Lycée à la Biochimie à l'Université
-
تحميل ملفات في مواضيع البيوكيمياء
Téléchargement de fichiers en Biochimie

Afin de pouvoir continuer à servir les visiteurs, soutenez nos actions sur le site takween en faisant acquisition des ouvrages et supports pédagogiques desitinés à améliorer l'enseignement et la recherche scientfique en Biochimie.


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com
{kind=link}