
La
peroxydase est une enzymes qui catalyse l'oxydation de molécules donneurs de protons H+ (comme les composés phénoliques des plantes) en présence du peroxyde d'hydrogène (eau oxygénée, H2O2) qui sera réduit en molécules d'eau (H2O).
L'étude de lacinétique d'une peroxydase (objet du travail pratique, TP) permet de déterminer le mécanisme de catalyse de cette réaction d'oxydoréduction complexe.
La deuxième partie du TP sera consacrée à l'étude de la structure 3D de la peroxydase du raifort .
Liens utiles:
- Vidéo test d'activité de la peroxydase
- Intérrogation
(début TP)
- Compte rendu TP peroxydases
| 1. INTRODUCTION GENERALE. Les peroxydases. Principaux types de mécanismes de catalyse des réactions enzymatiques. Technique d'électrophorèse. 2.MATERIEL ET METHODES. - Culture des plantes - Préparation du matériel végétal. - Extraction des peroxydases. - Fractionnement par le sulfate d'ammonium. - Chromatographie d'exclusion sur Séphadex G 100. - Dosage de l'activité enzymatique. |
3. DETERMINATION DU MECANISME DE CATALYSE DES PEROXYDASES. Cinétiques en fonction du gaiacol. Cinétique en fonction du peroxyde d'hydrogène. 4. SEPARATION DES ISOENZYMES DES PEROXYDASES. - Préparation des gels de polyacrylamide. - Electrophorèse. - Révélation des isoenzymes. 5.RESULTATS ET DISCUSSION. |
1. Introduction générale
Les
peroxydases sont des oxydoréductases qui catalysent l'oxydation d'un substrat (donneur de protons) en presence du peroxyde d'hydrogène (H2O2). La réaction globale est du type :
2 AH2 + H2O2 --------> 2 H2O + 2 AH.
Les radicaux libres formés (AH.) sont très réactifs et peuvent aboutir à la formation de polymères :
2 AH. A + AH2 (ou AH-AH)

Test de l'activité enzymatique de la peroxydase
En utilisant le gaiacol comme substrat (donneur de proton H+), la réaction donne naissance au tétragaiacol, composé rouge rouille (absorbant la lumière à 470 nm, environ.
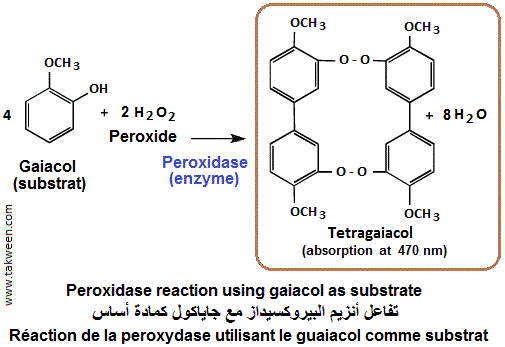
Aussi, l'o-dianisidine peut être utilisée comme substrat synthétique de la peroxydase
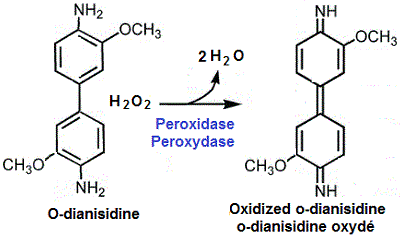
Les
peroxydases sont des hémoprotéines contenant
une composante glucidique. Leur groupement prosthétique est
constitué par la protohématine IX qui est une porphyrine
(4 noyaux pyrrols en cercle) liée à un atome de fer
sous forme ferrique.
Le fer peut contracter 2 autres liaisons (6 au total); une qui le
lie à l'azote d'une histidine au niveau de la partie protéique
et une autre liaison qui le lie à l'oxygène, lequel
peut faire partie d'une molécule d'eau, d'une molécule
d'oxygène, d'un peroxyde, etc.
Le noyau pyrrol est un hétérocycle pentagonal. Les multiples
doubles liaisons du groupement tetrapyrrolique sont conjuguées,
ce qui explique le pouvoir d'absorption de la lumière par la
porphyrine. Ainsi, les protéines à porphyrines sont
appelées chromoprotéines et sont toutes colorées
en rouge (peroxydases, hémoglobines,
cytochromes). Elles absorbent certaines radiations du spectre solaire
dont une étroite bande dans le proche ultraviolet (bande de
soret). La spécificité de leurs spectres d'absorption
est en rapport avec la structure moléculaire du groupement
prosthétique. L'étude spectroscopique permet de les
déceler, même à l'état de traces ou de
distinguer des molécules très voisines.
Dans le domaine des applications, les peroxydases peuvent être
utilisées dans plusieurs finalités dont les traitement
des eaux usées. En effet, elles permettent d'éliminer
les phénols (polluants toxiques) en catalysant leur polymérisation
(composés moins toxiques).
Les peroxydases purifiées à partir de nombreuses plantes
sont constituées par une seule chaîne polypeptidique.
Elles ont des poids moléculaires de 30000 à 50000. On
peut les trouver dans un même tissu sous forme de nombreuses
isoenzymes qu'il est facile de séparer et de purifier par
électrophorèse
en conditions non dénaturantes et par chromatographie d'échange
d'ions.
Principaux types de mécanismes de catalyse des réactions enzymatiques à 2 substrats.
Les
mécanismes
de catalyse des réactions enzymatiques à 2 substrats
sont de deux grands types :
- Les mécanismes où l'ordre de fixation des substrats
(S) et de libération des produits (P) ne compte plus. Ils sont
appelés mécanismes
au hasard (ou aléatoires).
- Les mécanismes où l'ordre de fixation des substrats
et de libération des produits est obligatoire. Ce sont des
mécanismes ordonnés ou séquencés.
- Les mécanismes n'impliquant pas de complexes ternaires et
forment une enzyme transitoire: mécanismes
ping-pong.
La cinétique enzymatique reste un moyen nécessaire mais
non suffisant, pour identifier les mécanismes de catalyse.
L'expérimentateur fait souvent recours aux modes
d'inhibition exercée par les analogues structuraux de l'un
des substrats vis à vis des autres substrats ou les types d'inhibition
manifestés par les produits de la réaction vis à
vis des différents substrats.
Les folioles du palmier dattier sont récupérées, lavées, numérotées et gardées au froid
3. Extraction et dosage des peroxydases
Prélever
5 g de tiges (partie entre la feuille et les racines) à partir
des jeunes plants de blé ou d'orge (selon les groupes). Après
rinçage avec l'eau distillée, broyer le matériel
végétal dans un mortier maintenu dans la glace, en présence
de 30 ml de tampon d'extraction :
- Acétate-acide acétique (pH 5,0) ......... 10 mM
- MgCl2 ………….............. 2 mM
- 2 mercaptoéthanol ……. 2 mM
- PVPP …………………. 5% (P/P)
Le 2 mercaptoéthanol empêche partiellement l'oxydation
des phénols en quinones qui peuvent provoquer l'inactivation
des enzymes.
Le PVPP, utilisé en milieu acide, adsorbe les composés
phénoliques susceptibles d'entraîner l'inactivation des
peroxydases.
L'homogénat obtenu est transvasé dans un tube de centrifugation
(pour rotor Sorvall SS34) puis centrifugé 10 min à 10000
g. Après centrifugation, le surnageant (extrait brut) est récupéré
dans une éprouvette de 100 ml. On note son volume final et
on en prélève un volume de 1,5 ml qui servira dans les
analyses suivantes.
Fractionnement par le sulfate d'ammonium.
Les protéines de l'extrait brut sont précipitées progressivement à froid, par le sulfate d'ammonium jusqu'à l'obtention d'un taux de saturation de 80% (51,6 g/100 ml). Après 2 heures et 30 min de précipitation à 0°C-4°C (glace) sous agitation lente, l'extrait est centrifugé à 10000 g pendant 10 min. Le culot obtenu est repris dans un faible volume (2 ml) de tampon acétate-acide acétique 10 mM (pH 5,0).
Chromatographie d'exclusion sur Sephadex G100.
L'extrait protéique obtenu par précipitation à 80% de saturation en sulfate d'ammonium, est filtré sur une colonne (1 cm x 15 cm) remplie de Sephadex G100. Après dépôt de l'extrait, 12 fractions de 1,5 ml sont collectées en bas de la colonne (chromatographie d'exclusion basée su le critère de la taille des molécules). Le dosage de l'activité enzymatique sera effectué sur l'ensemble des fractions. Il permet de tracer le profil d'élution des peroxydases. Les 2 fractions les plus actives du/des pic(s) seront rassemblées, séparément et conservées pour l'évaluation de la purification (rendements, taux de purification,…).
Dosage de l'activité enzymatique des peroxydases.
L'activité enzymatique est déterminée par spectrophotométrie
(voir vidéo spectrophotometrie
dans takween supports) en utilisant le gaiacol et le peroxyde
d'hydrogène (H2O2) comme substrats de l'enzyme.
Dans une cuve de 1 ml, déposer, dans l'ordre, 0,5 ml
de tampon acétate 0,1 M (pH 5,0), 0,4 ml de gaiacol
(à une concentration précisée selon le test,
voir tableau ci dessous), 0,050 ml d'enzyme et 0,050
ml de H2O2 (à une concentration précisée
selon le test) qui servira pour déclencher la réaction.
Le réglage à zéro de la densité optique
(DO) est réalisé après ajout de l'enzyme.
Après déclenchement de la réaction par addition
de H2O2 (concentration déterminée), la cuve, fermée
à l'aide d'un papier aluminium, est agitée rapidement
(par inversion dans les deux sens), puis introduite dans le spectrophotomètre,
préalablement réglé sur une longueur d'onde égale
à 470 nm. De suite, le temps de la réaction est suivi
à l'aide d'un chronomètre. Lire la DOà 470
nm au bout de chaque minute pendant une durée de 3 minutes.
Noter la variation de DO entre la la 3ième et la 2ème
et minute et entre la 2ième et la 1ière minute. Une
unité enzymatique correspond à la quantité de
protéines donnant 0,1 unité de DO à 470 nm.
- Vidéo: test de l'activité peoxydase chez le palmier dattier :



Détermination du mécanisme de catalyse des peroxydases
A partir de 4 concentrations de gaiacol et 4 concentrations de H2O2, faire les cinétiques pour chaque couple de concentration représenté dans le tableau ci-dessous et calculer les variations de DO correspondantes:
| Solution mère de H2O2 (%)* | Solution mère de gaiacol (mM) | |||
| 2 | 4 | 8 | 16 | |
| 0,04 | ||||
| 0,08 | ||||
| 0,16 | ||||
| 0,32 | ||||
* H2O2 1% présente une DO à 240 nm égale à 0,045 (Coefficient d'extinction de H2O2 à 240 nm = 43,6/M.cm).
Séparation des isoenzymes des peroxydases par électrophorèse non dénaturante sur gel de polyacrylamide
L'électrophorèse est faite verticalement sur des gels en polyacrylamide (voir vidéo de l'éléctrophorèse non dénaturante des isoenzymes, cas du palmier dattier).
Electrophorèse
L'électrophorèse est une méthode d'analyse et de séparation basée sur les critères de la charge électrique et la taille
des molécules. La migration différentielle de particules
chargées électriquement, se fait sous l'influence d'un
champs électrique. Seules les particules chargées positivement
ou négativement sont attirées par les pôles opposés
du champs électrique. Les composés qui peuvent être
transformés en particules chargées par formation de
complexes, sont de même sujets à une migration sous l'effet
du champs électrique.
Le pH des solutions joue un rôle très important dans
l'acquisition des charges électriques.
Ainsi, dans le cas des protéines, le pH auquel la charge électrique
globale est nulle est appelé point isoélectrique (PI).
L'électrophorèse peut être réalisée dans différentes conditions
dont celles qui ne dénature pa l'enzyme (conditions non dénaturantes).
Liens utiles:
- QCM-Electrophorèse. Supports de migration
- QCM-Electrophorèse (aspects techniques)e
Préparation des gels de polyacrylamide.
Les gels de polyacrylamides ont une épaisseur de 1 mm et sont confectionnés à l'aide de 2 plaques de verre de dimension 18 X 14 cm. La partie supérieure du gel, lieu du dépôt des extraits enzymatiques, présente une texture à porosité large (gel de concentration, 5% en polyacrylamide). Ce gel permet de concentrer les extraits avant leur séparation sur le gel à porosité étroite (gel de séparation, 10% en polyacrylamide).
La composition des 2 gels est résumée dans le tableau suivant :
| Gel de Concentration 5% (10ml) | Gel de séparation 9% (20 ml) | |
| Acrylamide-bis acrylamide 30%/0,8% (ml) | 1,70 | 6,70 |
| Tris-HCl 3M (pH 8,8) (ml) | - | 1,78 |
| Tris-HCl 0,5M (pH 6,8) (ml) | 2,5 | - |
| Eau distillée (ml) | 5,29 | 11,00 |
| Persulfate d'ammonium 1,5% (ml) | 0,50 | 0,50 |
| TEMED (ml) | 0,01 | 0,02 |
Electrophorèse.
Des volumes de 50 µl des différents extraits, préalablement mélangés avec 20 µl d'une solution de bleu de bromophénol, sont déposés dans les puits du gel de concentration. L'électrophorèse est faite à 35 mA. Le tampon d'électrodes (pH 8,3) est constitué de Tris 0,025 M, Glycine 0,192 M. Elle dure 5heures, environ (elle est arrêtée lorsque le bleu de bromophénol est à 0,5 cm de l'extrémité inférieure du gel).
Révélation des isoperoxydases.
Du gaiacol 0,023 M est préparé dans un tampon acétate 0,1 M (pH 5,0). Le gel ayant servi de support d'électrophorèse est trempé dans cette solution pendant 5 min. La révélation des peroxydases est ensuite déclenchée par addition de 1 ml de H2O2 1%. Laisser réagir 10 min à 40°C puis rincer le gel avec l'eau.
Résultats et discussion
Purification des peroxydases et détermination du mécanisme de catalyse.
* Tracer le profil d'élution des peroxydases.
* Déterminer les activités peroxydasiques de l'extrait brut et des fractions
enzymatiques obtenues par assemblage des contenus des tubes (pic(s) d'activité) après chromatographie sur Sephadex
G100. Calculer les rendements de purification pour chaque étape.
* Calculer les concentrations finales (en mM) du peroxyde d'hydrogène et du gaiacol utilisées dans la détermination du mécanisme de catalyse des peroxydases.
* Pour chaque fraction enzymatique obtenue après filtration sur
Sephadex G100, donner les tableaux (à double entrées) des activités (en variation de DO à 470 nm) déterminées en présence des différentes concentrations (en mM) de gaiacol et de peroxyde d'hydrogène.
* Tracer les diagrammes primaires 1/v = f(1/[S]) pour chacun des 2 substrats.
En déduire les diagrammes secondaires.
* Proposer le(s) mécanisme(s) susceptible(s) de rendre compte
des cinétiques obtenues.
* Calculer les valeurs des paramètres cinétiques Km et
Vmax. Les comparer à celles trouvées par le Logiciel Leonora.
* Donner le schéma de Cleland correspondant au mécanisme de catalyse des peroxydases.
Séparation par électrophorèse des isoperoxydases.
* Comparer la méthode de séparation des isoperoxydases par électrophorèse à celle effectuée à l'aide du Sephadex G100.
* Faire un schéma du zymogramme obtenu et mesurer les Rf des différentes bandes.
* Comparer les isoperoxydases de l'espèce végétale étudiée avec celles des autres espèces étudiées conjointement.
* Interpréter les résultats obtenus.
Partie 2. Peroxydase. Etude de la relation structure-fonction par visualisation tridimensionnelle à l'aide des programme Rastop et JMOL
La partie 2 du TP sera consacrée à l'étude de la structure 3D de la peroxydase du raifort (Horseradish, فجل) par le programme RasTop afin de visualiser les structures enzymatiques dont le site actif de l'enzyme.
QUESTIONS A REPONSE VIA LE PROGRAMME RASTOP:
Question 1.
Concernant les structures enzymatiques des peroxydases des plantes telle que la peroxydase du raifort, donner en s'aidant du programme RasTop une meilleure représentation graphique illustrant l'existence d'une molécule d'eau entre l'histidine distale (His42) de la zone polaire et le fer de l'hème.
Question 2. Concernant les structures enzymatiques des peroxydases des plantes telle que la peroxydase du raifort, donner en s'aidant du programme RasTop une meilleure représentation graphique illustrant la nature hydrphobe de la zone apolaire de l'environnement de l'hème.
Question 3. Concernant les structures enzymatiques des peroxydases des plantes telle que la peroxydase du raifort, donner en s'aidant du programme RasTop une meilleure représentation graphique illustrant le point d'attachement de l'hème à l'apoprotéine de l'enzyme.
Question 4.Concernant les structures enzymatiques des peroxydases des plantes telle que la peroxydase du raifort, donner en s'aidant du programme RasTop une meilleure représentation graphique illustrant l'état hexacoordonné de l'atome de fer de l'hème.
Sauver le travail sous forme d'un fichier script portant un nom indiquant le groupe de TP et les noms des deux étudiants ayant participé au travail (exemple 3-hilal-hanafi à placer dans le répertoire
LIENS VERS LA PARTIE 2 DU TP:
- PARTIE 2 (ETUDE DE LA STRUCTURE DES PEROXYDASES)
- Programme Rastop
LIENS UTILES
- -
Peroxydases du cactus (Opuuntia ficus indica L.)
- Peroxydases du figuier de barbarie
- Peroxydases de céréales
- Peroxydases des palmiersdattiers mâles et femelles
- Peroxydases des plantes. Théorie et pratique (Conférence
de Baaziz, 2006)
- - Etude de la structure 3D de la peroxydase

Chaine
Youtube (abonnement). Vidéos multilingues

- تحميل ملفات في مواضيع البيوكيمياء
Téléchargement de fichiers en Biochimie


Faculty of Sciences, Cadi Ayyad University
Marrakech, 40000, Morocco
Email: baaziz@uca.ac.ma
Phone: 212524434649 (post 513)
Fax: 212524434669
About takween.com
